Adjusted models for individual culm biomass of guadua bamboo (Guadua angustifolia Kunth) in managed stands in Southern Costa Rica
Modelos ajustados para biomasa de culmos de guadus (Guadua angustifolia Kunth) individuales en rodales manejados en el su de Costa Rica
Elemer Briceño-Elizondo1
Fecha de recepción: 13 de julio de 2018
Fecha de aprobación: 9 de octubre de 2018
Briceño-Elizondo, E. Adjusted models for individual culm biomass of guadua bamboo (Guadua angustifolia Kunth) in managed stands in Southern Costa Rica. Tecnología en Marcha. Vol. 32-2. Abril-Junio 2019. Pág 3-17.
DOI: https://doi.org/10.18845/tm.v32i2.4345
Keywords
Guadua angustifolia Kunth; carbon; biomass; management; forest modelling.
Abstract
This study aims to adjust field data to non-lineal models in order to predict growth and estimate biomass accumulation according to allometric data of managed bamboo plantations from productive sites. The plantation is located near the community of Rio Claro in the district of Guaycará, Canton of Golfito, Province of Puntarenas, Costa Rica. Five stands were identified for the study, with three of them used for calibration and two for model validation. A classification by height cohort was needed in order to organize the database for better model adjustment; namely H1:0 to 4m., H2: 4.01 to 7m., H3: 7.01 to 10 m., H4: higher than 10.01 m. The inventory data was fit through non-lineal regression to three selected biomass models used in previous bamboo biomass studies. The predicting variables used were diameter, height and basal area.
The quality of model prediction was tested sing a Wilcoxon test for paired samples; the empirical biomass from the validation data set was paired to model predictions from the three models. Predictions from model Bm1, Bm2 and Bm3 are not significantly different from empirical data (p= 0.7064, 0.7735 0.8822 respectively). Only Bm1 tends to overestimate but the range is acceptable for the prediction. The average results here obtained 12.49, 12.82 and 12.9 Mg ha-1 of biomass for Bm1, Bm2 and Bm3 respectively, are a lower to those found by other authors in Moso bamboo forest of southern China where they found 8.13±2.15 Mg ha-1 yr-1. Although the comparison is against another species and different maximum age of the plantation (5 years in Moso compared to 3 years) it can be suggested that, as in trees, a range of biomass according to age, management and dimensions is expected.
Resumen
Este estudio tiene como objetivo ajustar datos de campo a modelos no lineares con el fin de predecir el crecimiento y estimar la acumulación de biomasa en plantaciones de bambú bajo manejo en sitios productivos. La plantación está ubicada cerca de la comunidad de Rio Claro en el distrito de Guaycará, Cantón de Golfito, Provincia de Puntarenas, Costa Rica. Cinco rodales fueron identificados para el estudio, con tres de ellos utilizados para la calibración y dos para la validación de modelos. Una estratificación por cohorte de altura fue necesaria para organizar la base de datos a un mejor ajuste; a saber H1: 0 a 4 m., H2: 4,01 a 7 m., H3: 7,01 a 10 m., H4: superior a 10,01 m. Los datos de inventario se ajustaron mediante regresión no lineal a tres modelos de biomasa seleccionados utilizados en estudios anteriores de biomasa de bambú. Las variables predictivas utilizadas fueron diámetro, altura y área basal.
La calidad de la predicción del modelo se probó mediante la prueba de Wilcoxon para muestras pareadas; la biomasa empírica de la base de datos usada en validación se emparejó a las predicciones modelo de los tres modelos. Las predicciones de los modelos Bm1, Bm2 y Bm3 no son significativamente diferentes de los datos empíricos (p = 0.7064, 0.7735 0.8822 respectivamente). Sólo Bm1 tiende a sobrestimar, pero el rango es aceptable para la predicción.
Los resultados promedio obtenidos aquí son 12,49, 12,82 y 12,9 Mg ha-1 de biomasa en los modelos Bm1, Bm2 y Bm3 respectivamente, son mayores a los encontrados por otros autores en bosques de bambú Moso en el sur de China donde encontraron 8,13 ± 2,15 Mg ha-1 Año-1. Aunque la comparación es con otra especie y la edad máxima es diferente de la plantación (5 años en Moso en comparación con 3 años) se puede sugerir que, al igual que en árboles, se espera un rango de biomasa según edad, manejo y dimensiones.
Introduction
The renewed interest in the establishment, management, use and trade of guadua bamboo (Guadua angustifolia Kunth) for construction and other uses has led to a revival in interest in this natural resource, at a higher level than previously achieved. The genus Guadua spp is the most representative with the one with the greatest impact on human activities, since it brings together the largest and most economically important species of tropical America. This genus has more than thirty species, which are distributed from Mexico to Argentina, and are found from low elevations up to 2000 m. Its geographic distribution is mainly related to temperature, since it does not tolerate prolonged periods of very cold temperatures [1].
In Costa Rica, the most successful crops of the Guadua genus are between 240 and 500 m altitude, in areas with average annual rainfall of 3000- 4000 mm. It is difficult to determine the precise origin of the species and variations of guadua present in Costa Rica. It is suspected that some were directly imported from Colombia, Brazil and Peru [2]. As Montiel and Murillo [3] indicate, very particular morphological variations, identify the variations locally known as “South” due to being planted mostly in the southern region and “Atlantic” due to the same reason. There is a palpable development of the species in the Atlantic region as well as the South region of Costa Rica. Other areas have presence of guadua and other bamboo species, but not to the commercial avail found on these other two areas of the country. The level of organization varies from region to region as well; the south region shows successful enterprises working in the furniture industry, added value products, and culm length sales [4], as well as organized cooperatives planting the species along other crops in hopes of a potential market [5]. Many producers have been also encouraged to establish plantations within the scope supplying raw material to new potential products with high added value [6].
Once full dimensions are achieved, the bamboo culms enter a period of fiber maturity which can last up to 7 years depending on the species, after which they deteriorate rapidly, releasing carbon back into the atmosphere [7]. Therefore in a natural state, bamboo will reach a stable level of above ground carbon relatively quickly, where carbon accumulation through sequestration is counter off when decay comes. In order for the bamboo system to continue to be a net sink, carbon has to be stored in other forms, so that the total accumulation of carbon in a solid state exceeds the carbon released to the atmosphere. Chapters 7 and 8 discuss these questions, amongst other issues that can affect the length of storage of carbon [8].
In several studies, generally, at ages between five and seven years, a high variability of culms per hectare is reported; for example, from 5090 to 9416 culms ha-1 according to Arango-Arango and Camargo [9], 11 827 ± 3884 culms ha-1; according to Camargo-García, Rodriguez y Arango [10], 4050 culms ha-1 and 10101 culms ha-1 in the work of Castañeda-Mendoza, Vargas-Hernandez, Gomez-Guerrero, Valdez-Hernandez, Vaquera-Huerta [11]. The objective of this study is to adjust models of bamboo growth in order to predict growth and estimate biomass accumulation according to allometric data and the potential for carbon sink of managed bamboo plantations in a productive site in southern Costa Rica.
Material and methods
Study area
The selected plantation (8°71’N, -83°09’O) is located near the community of Rio Claro in the district of Guaycará, Canton of Golfito, Province of Puntarenas, Costa Rica (figure 1). The site is property of a bamboo producing company with a large trajectory of bamboo planting, silviculture and commerce. Elevation is between 200 and 300 meters above sea level; the annual rainfall is between 4000 to 5000 mm with an average temperature of 25°C. The bamboo area has different strata, which are differentiated by topography and age; with an area of 11.74 ha total. The property includes other vegetation coverage, classified official as non-forest land, forest and part of secondary forest. Soils are classified as entisols and inceptisols (suborder orthens and udepts).
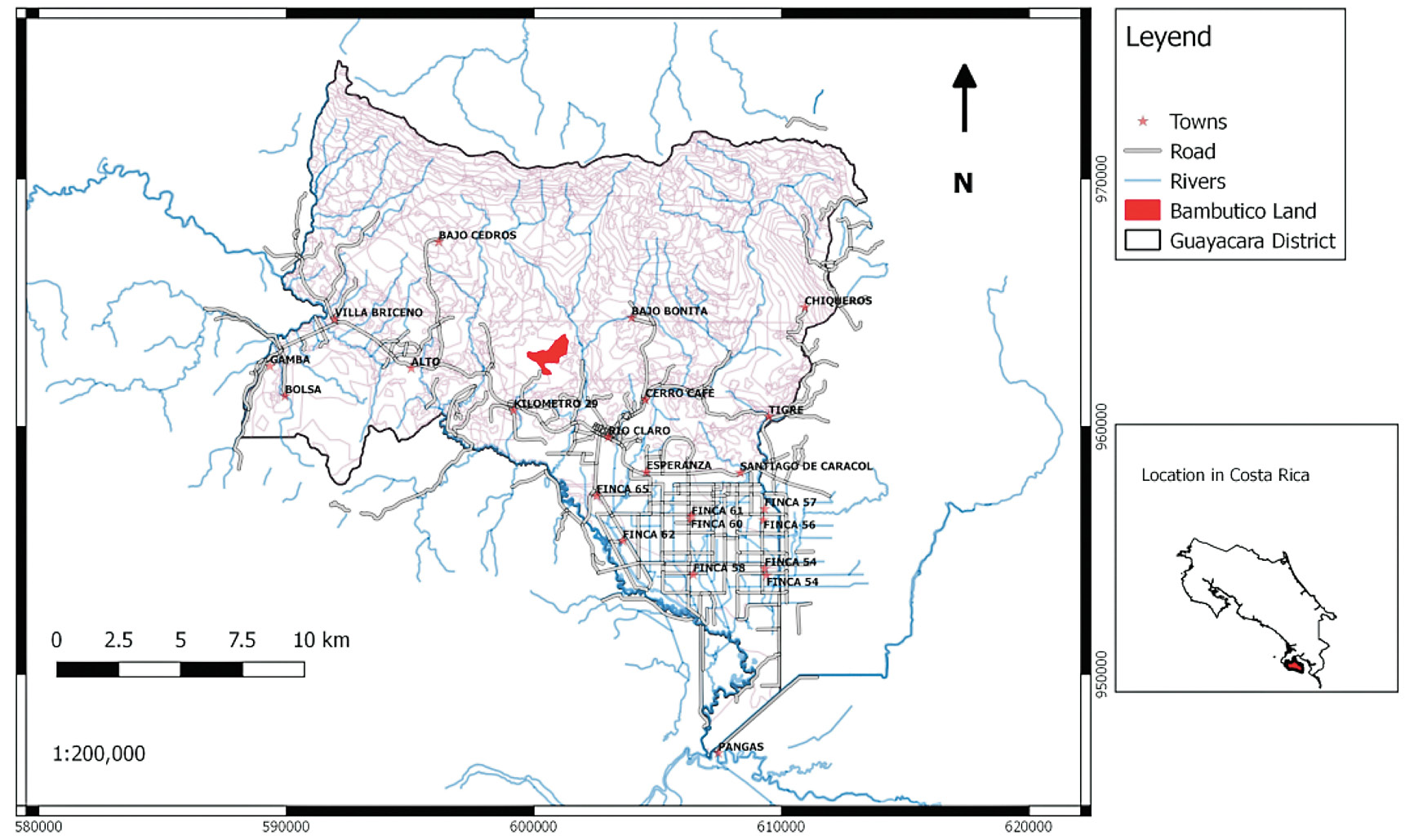
Figure 1. Site location within the Guaycará district of the 3 year old guadua bamboo plantation.
The terrain presents an evident deposition of organic matter, rocky outcropping all around the stand, but well planned and maintained inner roads.
Management regime
The site received intensive and meticulous management; the silvicultural practices include early elimination of primordial shoots, thinning of small culms, and pruning and bud control. The stand is well established, and despite its young age (3 years) already has a canopy closure (figure 2). These practices took place after the first year of establishment to guarantee that the clump has already establish itself ion the site. Normally only commercial shoots are left to reach 11± 4 culms in average at each clump. Thinning was designed to maintain a desired culm density per hectare, the average culm density of the plantation is 1942 culms ranging from 1105 culms in one strata up to 3166 in the denser strata. Thinning schedule and intensity followed recommendations based upon inventory data for 2015. Initial weed control takes place at establishment with a proper weeding for each individual, followed by manual control until canopy closure. Bud control consists of the mechanical impediment of branch expansion, where a light punch is applied to the branch bud before the branch’s emergence, facilitating later handling of the culm and saving management costs. Not all branches are treated as such due to the need of a healthy canopy, thus pruning is necessary before harvest, in order to facilitate extraction.


Figure 2. Managed Guadua angustifolia clump within Guaycará plantation of three years of age.
Inventory Data (2015-2016)
The sampling method was a modification of triplet sampling method for individual trees [12], renamed here “Individual Clump Method” (ICM). Five stands were identified for the study within the plantation (table 1), with three of them used for calibration (1, 4, and 7) of the models and two for model validation (5, 6). The separation of the stands was based purely on administrative boundaries within the plantation, following slope arrangements, the previous facilitated management activities and served as a way of stratification. All stands had the same age and received the same management. The sampling unit is made up of a clump with all its culms. The initial clump is randomly chosen, and then according to the defined sampling intensity sampling continues up to the next fixed determined clump. For a 2% sampling intensity, a clump is chosen each 50 clumps (100/2 = 50) and for a 4% a unit is selected every 25 clumps (100/4 = 25). Once at the selected clump, all culms are measured taking information on culm diameter, height, state of maturity, and sanitary state. The number of culms measured in this method is equivalent to the establishment of a plot of 500 m2 where culms are randomly distributed throughout the stand [12].
The difference of this method being applied to a bamboo plantation from a tree plantation lies in the fact that the point of measurement yields data from many individual culms, allowing to capture more variation and information. This method is applicable to a plantation that has not lost its clump differentiation and is small in size. The method requires to survey the entire stand from start to finish by foot. Since all clumps in the plantation or stand are counted, it is possible to estimate the total number of culms and at the same time their state of maturity, which can then be extrapolated to hectare, namely:
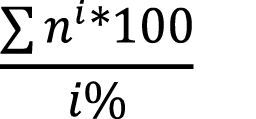 (equation 1)
(equation 1)
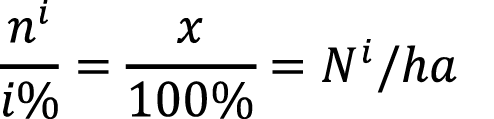 (equation 2)
(equation 2)
Where: ni: total culms in the clump or per state of maturity
i%: sampling intensity.
For the determination of empirical biomass and carbon, data from the inventory was used in combination with reported factors from literature for wall thickness, tapper and bamboo wood density [13]. The volume of the cylinder was first used, then allometrically reduced using the tapper factor, to get apparent volume. This was reduced by taking into account the average wall thickness reported for the species [14], in order to eliminate the “empty” volume of the interior of the culm. The inner empty volume was subtracted to get real volume present for the bamboo culm. The information on the specific weight of the species [15] for conversion is used to get biomass of the culm. Other components are left aside since there is not reliable information and the rhizome and leave remain at the site.
 (equation 3)
(equation 3)
Where: B =empirical biomass of culm i within strata i (kg).
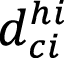 = diameter of culm i within strata i (cm).
= diameter of culm i within strata i (cm).
= height of culm i within strata i (m).
ffg=tapper factor for G.angustifolia.
Dkg= specific density in kg m-3.
A classification by height cohort was needed in order to organize the database for better model adjustment; namely H1:0 to 4m., H2: 4.01 to 7m., H3: 7.01 to 10 m., H4: higher than 10.01 m (table 1). The stratification of the database in these cohorts allows for a more precise model parameterization given than a single culm, according to the date of data collection will exhibit a height which will change in time, keeping however the same diameter from emergence to harvest. Three stands were used for model calibration and two for model validation.
Model selection and model adjustment.
The inventory data was fit through non-lineal regression to three selected biomass models (equations 4-6) used in previous bamboo biomass studies. The predicting variables used were diameter ([16]: equation 4), diameter and height ([17]: equation 5) and basal area ([18]: equation 6). The previous cohort stratification was declared in model adjustment for the calibration runs
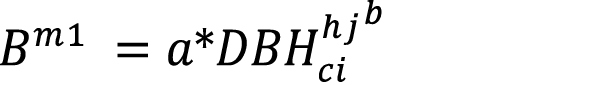 (equation 4)
(equation 4)
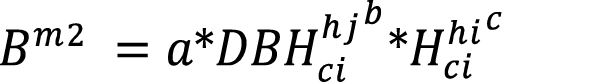 (equation 5)
(equation 5)
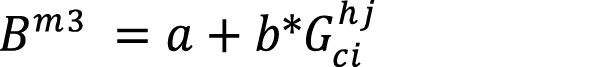 (equation 6)
(equation 6)
Where: 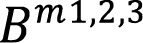 = Biomass model (kg).
= Biomass model (kg).
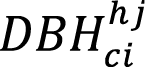 = diameter at breast height of culm i within height cohort j (cm).
= diameter at breast height of culm i within height cohort j (cm).
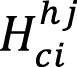 = height of culm i within height cohort j (m).
= height of culm i within height cohort j (m).
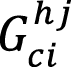 =basal area of culm i within height cohort j (m2).
=basal area of culm i within height cohort j (m2).
a,b,c= parameters.
Table 1. Mean DBH, height and stand characteristics for G.angustifolia plantation within Guaycará district used for model calibration.
|
Stand |
cohort |
Stage of Maturity |
DBH(cm) |
Height (m) |
|
1 |
1 |
J |
3,50±1,98 |
2,85±0,85 |
|
R |
10,13±2,06 |
2,98±0,79 |
||
|
2 |
J |
4,58±1,50 |
6,05±0,91 |
|
|
R |
10,98±1,11 |
6,08±0,90 |
||
|
3 |
J |
6,54±2,48 |
8,59±0,72 |
|
|
R |
10,23±1,67 |
9,00±1,00 |
||
|
4 |
J |
9,15±1,70 |
12,81±0,91 |
|
|
R |
10,65±1,45 |
11,50±0,97 |
||
|
4 |
1 |
J |
2,86±1,29 |
3,56±0,63 |
|
R |
10,15±2,37 |
2,69±0,63 |
||
|
2 |
J |
4,78±2,62 |
5,28±0,72 |
|
|
R |
11,17±1,43 |
5,71±0,95 |
||
|
3 |
J |
7,59±1,76 |
8,91±0,89 |
|
|
R |
9,45±1,04 |
9,17±0,75 |
||
|
4 |
J |
8,85±1,22 |
12,38±1,06 |
|
|
R |
8,90±0,83 |
12,00±0,82 |
||
|
7 |
1 |
J |
3,17±1,71 |
3,46±0,64 |
|
R |
9,86±1,67 |
3,38±0,74 |
||
|
2 |
J |
4,32±1,84 |
5,39±0,74 |
|
|
R |
9,30±1,70 |
5,50±0,71 |
||
|
3 |
J |
7,28±1,13 |
9,05±0,79 |
|
|
R |
8,10±0,00 |
8,00±0,00 |
||
|
4 |
J |
7,29±1,09 |
11,40±0,75 |
|
|
R |
10,00±0,00 |
13,00±0,00 |
Sum of strata and Carbon calculations
Data from the inventory, model validation runs were sum up by stand and extrapolated to hectare basis in order to give an estimate of the carbon reservoir present and carbon sequestration potential of a well-managed plantation. These data were compared to empirical biomass present in the calibration stand by means of an analysis of variance.
Results
Model runs
Inventory data was fit to the selected models to approximate its variables; the results are presented in table 2. Allometric non lineal models were fit to empirical biomass to approximate the constants for each model for each cohort. The p-values show significance for each constant used in the models (p< 0.05). The values were fit to respond to height cohorts and stage of maturity. An important difference among the models tested is the type of variable used, where in one of the models occupancy of the site takes relevance over individual dimension. The number of individuals to make sprout biomass predictions were less than for young culms due to the dynamics of bamboo stands, where it is expected to have less sprouts than young or mature culms at any age of the stand.
Table 2. Culm biomass estimation models for Guadua angustifolia Kunth, using non-lineal regression, for different stages of maturity and height cohorts. Biomass is expressed in kg
|
Model |
Growth stage |
Cohorts |
N |
Parameters |
||||||
|
a |
b |
c |
Cmerror |
p-value |
p-value |
p-value |
||||
|
(a) |
(b) |
(c) |
||||||||
|
Bm1 |
young |
1 |
103 |
0.28 |
1.31 |
--- |
0.22 |
<0.0001 |
<0.0001 |
--- |
|
2 |
140 |
0.49 |
1.31 |
--- |
0.5 |
<0.0001 |
<0.0001 |
--- |
||
|
3 |
67 |
0.89 |
1.24 |
--- |
1.11 |
<0.0001 |
<0.0001 |
--- |
||
|
4 |
59 |
1.2 |
1.26 |
--- |
1.96 |
<0.0001 |
<0.0001 |
--- |
||
|
Sprout |
1 |
24 |
0.25 |
1.32 |
--- |
1.85 |
0.2365 |
0.001 |
--- |
|
|
2 |
21 |
1.48 |
0.84 |
--- |
2.94 |
0.1776 |
0.0106 |
--- |
||
|
3 |
17 |
0.54 |
1.45 |
--- |
1.82 |
0.0115 |
<0.0001 |
--- |
||
|
4 |
15 |
1.57 |
1.1 |
--- |
3.1 |
0.0172 |
<0.0001 |
--- |
||
|
Bm2 |
young |
1 |
103 |
0.0645 |
1.4307 |
1.0642 |
0.0148 |
<0.0001 |
<0.0001 |
<0.0001 |
|
2 |
140 |
0.0827 |
1.2816 |
1.0527 |
0.0356 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
3 |
67 |
0.103 |
1.2003 |
1.02 |
0.0201 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
4 |
59 |
0.1161 |
1.1674 |
0.9983 |
0.0102 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
Sprout |
1 |
24 |
0.1206 |
1.1476 |
1.0014 |
0.0004 |
<0.0001 |
<0.0001 |
<0.0001 |
|
|
2 |
21 |
0.1262 |
1.1293 |
1.0002 |
0.0002 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
3 |
17 |
0.124 |
1.1408 |
0.9957 |
0.0011 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
4 |
15 |
0.1268 |
1.1293 |
0.9979 |
0.0017 |
<0.0001 |
<0.0001 |
<0.0001 |
||
|
Bm3 |
young |
1 |
103 |
0.45 |
798.83 |
--- |
0.32 |
<0.0001 |
<0.0001 |
--- |
|
2 |
140 |
1.32 |
1213.85 |
--- |
0.92 |
<0.0001 |
<0.0001 |
--- |
||
|
3 |
67 |
3.68 |
1519.77 |
--- |
1.68 |
<0.0001 |
<0.0001 |
--- |
||
|
4 |
59 |
6.46 |
1914.82 |
--- |
2.15 |
<0.0001 |
<0.0001 |
--- |
||
|
Sprout |
1 |
24 |
1.47 |
1914.82 |
--- |
1.89 |
0.1055 |
0.0002 |
--- |
|
|
2 |
21 |
6.22 |
507.04 |
--- |
2.95 |
0.0016 |
0.009 |
--- |
||
|
3 |
17 |
4.15 |
1408.14 |
--- |
1.78 |
0.0022 |
<0.0001 |
--- |
||
|
4 |
15 |
9.51 |
1305.53 |
--- |
3.04 |
0.0001 |
<0.0001 |
--- |
||
|
DBH: Diameter at breast height (cm); H: height (m); G: basal area (m2); young: young culm, Sprout: sprout culm Cohorts: (1) from 0 to 4 meters in height ,(2) from 4.01 to 7 meters in height, (3) from 7.01 to 10 meters in height, (4) higher than 10.01 meters in height; N: number of individuals; a,b,c: parameters; Bm1: Biomass (kg) = a*DBHb; Bm2: Biomass (kg)= a*DBHb*Hc; Bm3: Biomass (kg)= a+b*G. |
||||||||||
Validation
The stand characteristics of guadua bamboo used in the validation of the models is presented in table 3. The average DBH and height were divided into the height cohorts by stage of maturity, following the cohort separation used for calibration. The average DBH is higher in sprouts independently of the cohort, signaling that the new recruited culms have obtained the occupation capacity needed for bigger dimensions due to the absence of competition from older culms in the clump, influencing the new culms to reach higher dimensions; this is consistent with the correlation of mean DBH and stand density, where at lower density a higher DBH can be expected. The dimensions ranged from 2,87±1,07 up to 10,80±0,99 in stand 5 and from 2,78±1,26 up to 12,00±0,001 cm in stand 6. Some cohorts did not show data for sprouts due to aspects of presence in that height category.
Table 3. Mean DBH, height and stand characteristics for G.angustifolia plantation within Guaycará district used for model validation.
|
Stand |
cohort |
Stage of maturity |
DBH (cm) |
Height (m) |
|
5 |
1 |
Young |
2,87±1,07 |
3,83±0,16 |
|
Sprout |
9,20±0,001 |
4,00±0,001 |
||
|
2 |
Young |
5,31±2,46 |
6,68±0,68 |
|
|
Sprout |
8,47±1,10 |
5,67±0,58 |
||
|
3 |
Young |
7,11±2,18 |
8,30±1,05 |
|
|
Sprout |
10,80±0,99 |
8,50±0,71 |
||
|
4 |
Young |
8,14±1,52 |
12,00±0,001 |
|
|
Sprout |
0,00±0,001 |
0,00±0,001 |
||
|
6 |
1 |
Young |
2,78±1,26 |
3,54±0,54 |
|
Sprout |
9,70±0,82 |
3,50±0,50 |
||
|
2 |
Young |
5,91±2,77 |
6,18±0,87 |
|
|
Sprout |
0,00±0,00 |
0,00±0,00 |
||
|
3 |
Young |
7,45±0,98 |
8,67±1,03 |
|
|
Sprout |
11,60±0,57 |
8,50±0,71 |
||
|
4 |
Young |
8,10±0,00 |
11,00±0,001 |
|
|
Sprout |
12,00±0,00 |
11,00±0,001 |
||
|
DBH: Diameter at breast height (cm); H: height (m); G: basal area (m2); young: young culm, Sprout: sprout culm Cohorts: (1) from 0 to 4 meters in height ,(2) from 4.01 to 7 meters in height, (3) from 7.01 to 10 meters in height, (4) higher than 10.01 meters in height. |
||||
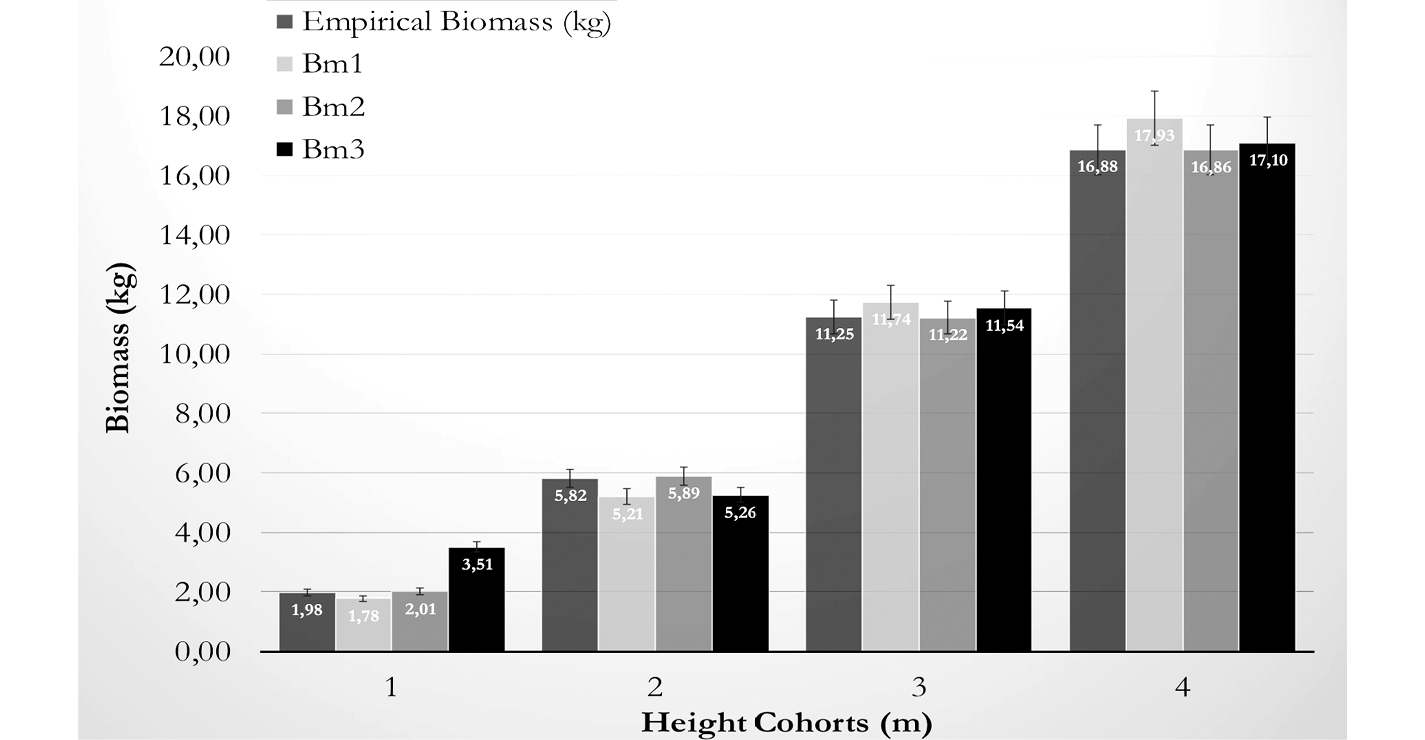
The quality of model prediction was tested sing a Wilcoxon test for paired samples; the empirical biomass from the validation data set was paired to model predictions from the three models. Table 4 shows that predictions from model Bm1, Bm2 and Bm3 are not significantly different from empirical data (p= 0.7064, 0.7735 0.8822 respectively). Predictions between models were also tested to realize differences on the number of variables or variables used; only Bm1 and Bm3 showed significance for their predictions (p: 0,0272).
Table 4. Wilcoxon test for paired samples, between empirical biomass (EB) and adjusted culm biomass models for G.angustifolia Kunth in the Guaycará site in Southern Costa Rica.
|
Obs(1) |
Obs(2) |
N |
Suma(R+) |
E(R+) |
Var(R+) |
DE(dif) |
p(2 tails) |
|
EB |
Bm1 |
87 |
2003 |
1914 |
55824,5 |
1,1 |
0,7064 |
|
EB |
Bm2 |
87 |
1982 |
1914 |
55824,5 |
0,14 |
0,7735 |
|
EB |
Bm3 |
87 |
1949 |
1914 |
55824,5 |
2,37 |
0,8822 |
|
Bm1 |
Bm2 |
87 |
1757 |
1914 |
55824,5 |
1,12 |
0,5064 |
|
Bm1 |
Bm3 |
87 |
2436 |
1914 |
55823,13 |
2,31 |
0,0272 |
|
Bm2 |
Bm3 |
87 |
2006 |
1914 |
55824,5 |
2,38 |
0,697 |
|
EB: empirical biomass, Bm1: Biomass model 1; Bm2: Biomass model 2; Bm3: Biomass model 3. |
|||||||
This difference in model prediction between Bm1 and Bm3 can be observed in figure 3, where individual average biomass per culm is shown. Estimations are also shown per cohort with the typical error for the prediction. Only Bm1 tends to overestimate but according to table 4 the range is acceptable for the prediction. The aim of model building is to have adequate culm biomass prediction which can be deducted from inventory data; the total biomass per hectare will depend on site density, which is a product of management.
Figure 3. Average biomass under three adjusted culm biomass models and empirical data for G.angustifolia in the Guaycará site.
The necessity to separate the models by cohort can be reflected on the behavior observed on figures 4 through 7 where it can be seen the agreements for the models with empirical data; model accuracy is better when the culm has finished its vertical growth as related to cohort 4.
Sum strata and extrapolation to hectare
The usefulness of the models however is finally realized when the values per hectare are calculated. The biomass per hectare can be seen in table 5, where it can be seen that the empirical biomass does not vary greatly when using the models to compile the amount of Mg ha-1 which consider also the actual density in the stands per maturity stage the biomass in stand. Stand 5 has a higher density than stand 6; where 8.7±0.15 Mg ha-1 of biomass are accumulated in empirical data and 8.55±0.15, 8.88±0.14, 8.35±0.14 Mg ha-1 of biomass are present when using Bm1, Bm2 and Bm3 respectively for 1185 young culms ha-1. The amount of biomass for young culms in stand 6 is lower than stand 5 as mentioned before, where the density reaches 985 culms ha-1; reporting 6.98±0.12 for empirical data and 6.89±0.11, 7.01±0.12, 6.72±0.11 Mg ha-1 of biomass for Bm1, Bm2 and Bm3 respectively. Sprout biomass has lower values due to the lower density of this maturity stage, where stand 5 and 6 have 173 and 148 culms ha-1 respectively; however, Bm3 shows higher biomass accumulation than the empirical data and the other two models.
Table 5. Mean empirical and modeled biomass (Mg ha-1), and mean culms ha-1 for G.angustifolia Kunth for G.angustifolia Kunth in the Guaycará site in Southern Costa Rica.
|
Stand |
Stage of maturity |
Culms ha-1 |
Biomass (Ton ha-1) |
|||
|
EB |
Bm1 |
Bm2 |
Bm3 |
|||
|
5 |
Young |
1185 |
8.79±0.15 |
8.55±0.15 |
8.88±0.14 |
8.35±0.14 |
|
Sprout |
173 |
3.93±0.12 |
3.94±0.13 |
3.94±0.12 |
4.55±0.10 |
|
|
6 |
Young |
985 |
6.98±0.12 |
6.89±0.11 |
7.01±0.12 |
6.72±0.11 |
|
Sprout |
148 |
3.54±0.13 |
3.70±0.15 |
3.56±0.13 |
5.26±0.12 |
|
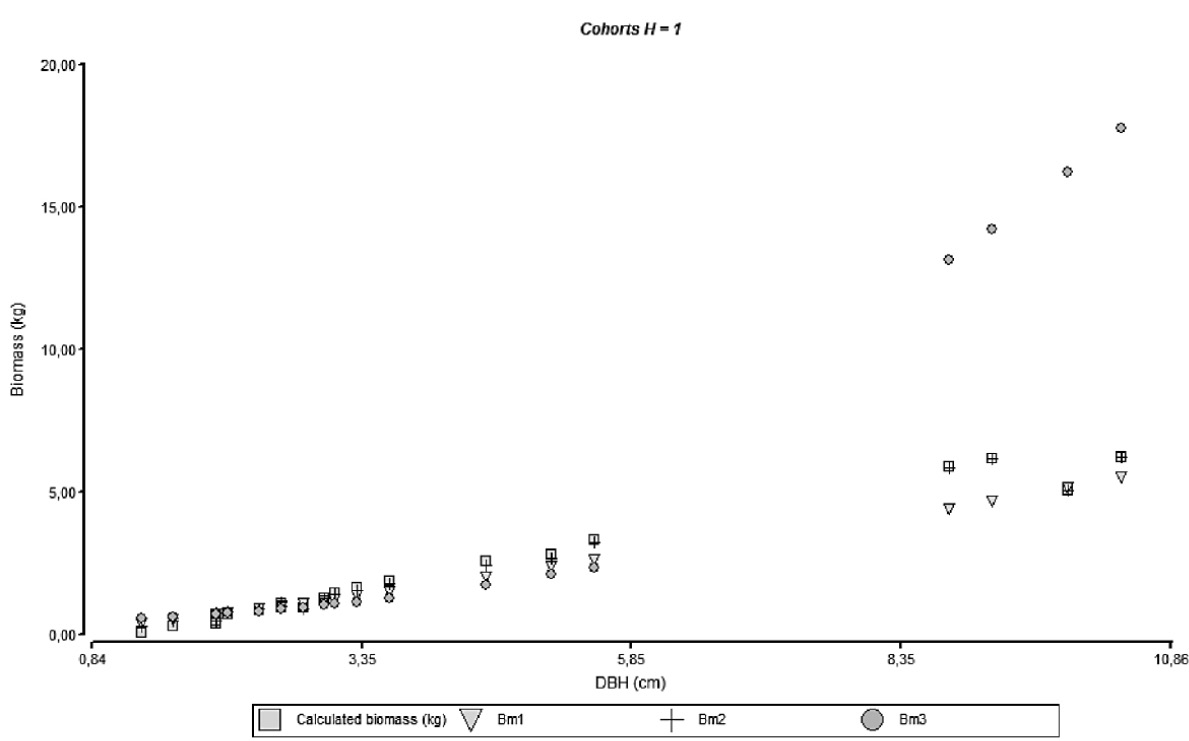
Figure 4. Relationship between diameter and biomass for the adjusted models and empirical data of different height cohorts.
Discussion
Biomass estimation is a step stone to quantify the amount of carbon locked into the biosphere. Such information aids on knowing the changes that occur when the forest is intervened, tracking changes in the carbon stocks of forest and as a way to fulfill international commitments on information on national carbon stocks. Direct method (destructive techniques) and indirect method (biomass equations) are generally used for biomass estimation of forests. Indirect methods such as biomass equations are less time consuming and more cost effective when compared to destructive methods, making them attractive tools in forest inventories. Different linear and non-linear models have been developed worldwide for estimating biomass, however due to the natural patterns of growth of most woody species, nonlinear equations are better suited for biomass estimation [19].
The present study aims at adjusting nonlinear equations to predict G.angustifolia culm biomass in managed stand for the species at a key site in southern Costa Rica. The selection of the models to use was based on previous studies on Moso bamboo, a very known and commercial species in Asia [16], [17] and a study base on allometry for G.angustifolia in northern Costa Rica [18]. There are many studies on allometric models for tree species, however bamboo seems neglected, despite being regarded as a woody plant; its growth patterns of woody bamboos are different from timber in that the former possess a woody vascular bundle structure of fast growth while the latter has clear tree rings [17]. Therefore, the use of models tested in some important species are of interest for G.angustifolia as well as models for the species which have been tested under different conditions, in order to project their usefulness with other management systems in the country.
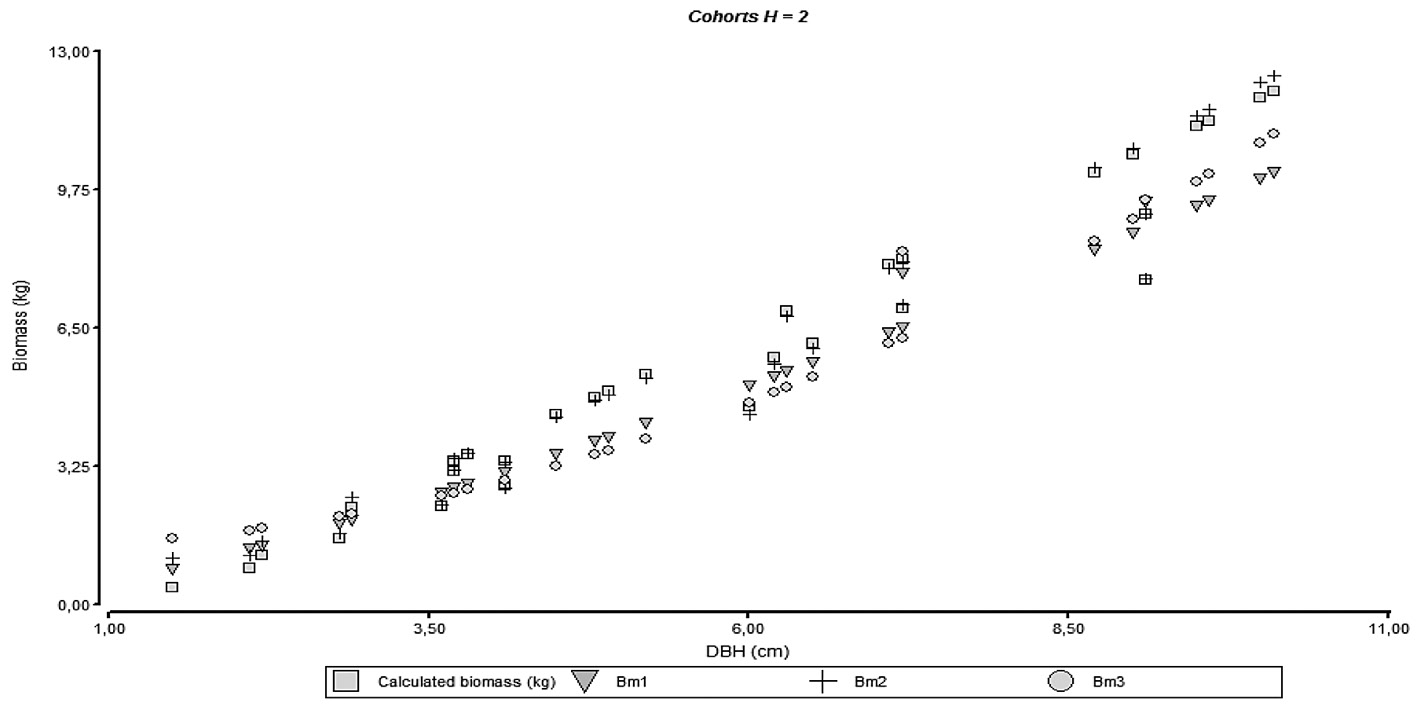
Figure 5. Relationship between diameter and biomass for the adjusted models and empirical data of different height cohorts.
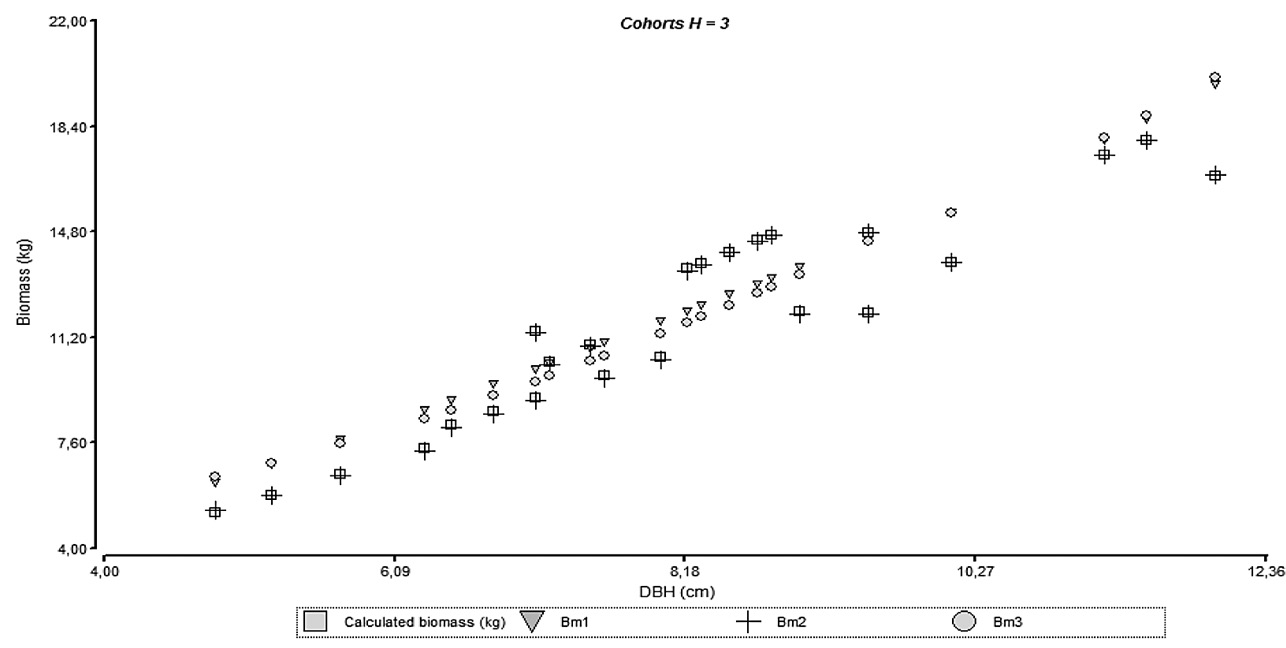
Figure 6. Relationship between diameter and biomass for the adjusted models and empirical data of different height cohorts
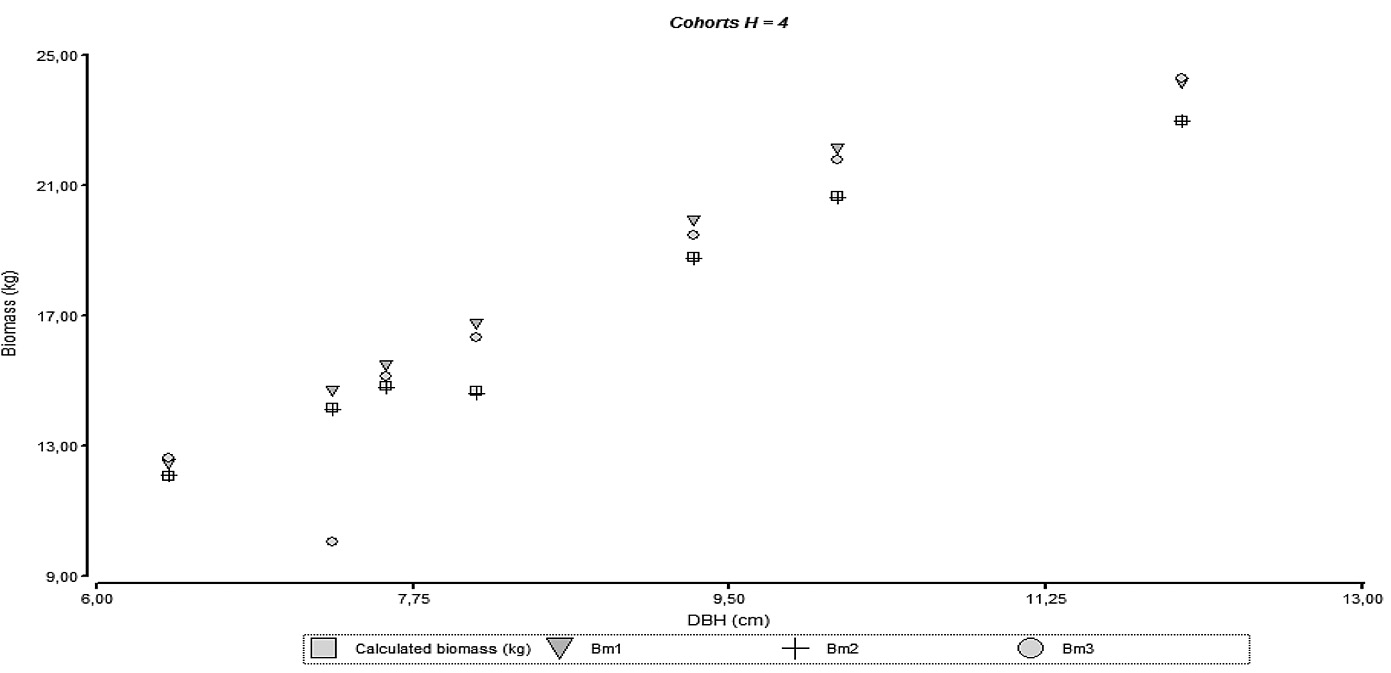
Figure 7. Relationship between diameter and biomass for the adjusted models and empirical data of different height cohorts.
The parameters for the models used depend on the nature of the data, therefore it was necessary to stratify the database to obtain a better fit of the model due to the dispersion of the data, as observed in the differences in DBH and height among the cohorts in the three stands used in calibration (table 1); equally this helps to better approximate model validity by focusing on a realistic number of culms that will endure until harvest or biological age is reached. The resulting stratification aided to obtain reliable biomass models for young culms; however, the model Bm1 cannot be reliable when predicting biomass for sprouts at low height cohorts, the p values obtain demonstrate that the models are sensitive to variable variations (table 2). The previous might be explained by the fact that the culm, has not yet reached total height, and the variation for height is too large; since Bm1 is base only on DBH for its prediction, it leaves this variable out when it can be seen that it can have an influence on the parameters and the prediction potential of the model. A similar situation is seen in Bm3 for the first cohort, where the model is based on basal area only. Other cohorts for this models seem to work adequately, whilst Bm2 includes height and is not subject to this source of variation. The models used in Yen et al [16], [17] are similar in terms of management and size as for the one suggested in Fonseca and Rojas [18] has similar size and weather conditions but no management; however, its age has taken it to a self-thinning stage which controls density.
Validation data had therefore similar dimensions, since all management in the site was standard, with sprouts reaching higher DBH dimensions. That pattern of growth in most bamboos would indicate that the plantation is increasing its biomass with time, by means of recruiting new culms with higher diameters. As table 4 indicates, there is no significance between empirical data and the models, except when comparing Bm1 and Bm3; the previous indicates that for all models tested (against empirical data), the predictions can be considered reliable, with the given degree of variation, which is answerable on the fact that two of the three models exclude height as a predicting variable, nevertheless the models are useful. The comparison between models aids to check congruency on the simulations, and the difference might arise from the combination of the entire data set when carrying on the comparison, as it must be remembered that the models carry different behaviors per cohort. The latest can be observed in figures 4 through 7, where observed and simulated data tend to be exponential at lower height cohorts to latter show a tendency to climax in the higher cohort. The use of DBH to predict biomass is a general method that has been widely applied in the bamboo forests [20]. This variable is the quickest and easiest available for allometry. In bamboo it is assumed that culm biomass accumulates mainly in the first year and then afterwards its biomass increased only slightly or not obviously compared with the same DBH; when other authors have used age, here a decision towards height stratification was followed to better approximate the levels of biomass variation, as the diameter within the same recruitment year might be similar if the same management is applied. This can be appreciated in figure 3, where the stratification shows a more accurate approximation for biomass.
The results here obtained (12.49 12.82 and 12.9 Mg ha-1 of biomass for Bm1, Bm2 and Bm3 respectively), are a lower to those found by Yen et al 2011 in Moso bamboo forest of southern China where they found 8.13±2.15 Mg ha-1 yr-1; although the comparison is against another species and different maximum age of the plantation (5 years in Moso compared to 3 years) it can be suggested that , as in trees, a range of biomass according to age, management and dimensions is expected. Management and soil conditions have an important role to play, the values in this study area subject to an intensive management which has its density from 1133 to 1358 culms ha-1 (table 5) as compared to 6500 to 7500 culms ha-1to that of Yen et al 2011, making them approximately 6 times lower in density than the Chinese bamboo forest. Studies in Costa Rica on plantations with high densities and cero management show values of 6 473± 10.8% culms ha-1 (all maturity stages) and 7.25 Mg ha-1 for sprouts and young culms. One could argue the necessity of management here, however, 42.03 Mg ha-1 are found within mature category [18], which did not get harvested, cramping growing space, and possibly possibilities to an optimized succession.
Allometric models are powerful tools that are widely applied to estimate volume, biomass and carbon storage of forests [16], [17]. As suggested by Zianis and Mencuccini [21], the most common variable used to predict volume, biomass and carbon storage is DBH. Bamboo plantations have a dynamic of growth different from that of trees and an effort to incorporate models that work with site occupancy and biomass are more reliable than volumetric models.
References
[1] X. Londoño, G. C. Camayo, N. M. Riaño, y Y. López, «Characterization of the anatomy of Guadua angustifolia (Poaceae: Bambusoideae) culms», Bamboo Science and Culture: The Journal of the American Bamboo Society, vol. 16, n.º 1, pp. 18–31, 2002.
[2] M. Montiel, V. M. Jiménez, y E. Guevara, “Caracterización anatómica ultraestructural de las variantes” Atlántica, Sur”, Revista de Biologia Tropical, vol. 54, n.º 2, pp. 1-12, 2006.
[3] M. Montiel Longhi y L. Murillo Rodríguez, “Historia ecológica y aprovechamiento del bambú”, Revista de biología tropical, vol. 46, pp. 11–18, 1998.
[4] Retana. Comunicacion personal. 2015.
[5] A. Arguedas-Chaverri, «Guadua angustifolia Kunth: opción de diversificación productiva para productores en la Península de Osa, Costa Rica.» Tesis de graduación. Escuela de Ingeniería Forestal, Instituto Tecnológico de Costa Rica, 2015.
[6] FONAFIFO. Comunicacion personal. 2017.
[7] W. Liese, “Bamboo as carbon sink-fact or fiction?”, Journal of Bamboo and Rattan, vol. 8, n.º 3/4, pp. 103–114, 2009.
[8] R. Düking, J. Gielis, y W. Liese, «Carbon flux and carbon stock in a bamboo stand and their relevance for mitigating climate change», J Am Bamboo Soc, vol. 24, n.º 1, pp. 1–7, 2011.
[9] A. M. Arango Arango y J. C. Camargo García, «Bosques de guadua del Eje Cafetero de Colombia: oportunidades para su inclusión en el mercado voluntario de carbono y en el Programa REDD+», Recursos Naturales y Ambiente. Número 61 (Diciembre 2010), páginas 77-85, 2010.
[10] J. C. Camargo, J. A. Rodríguez, y A. M. Arango, “Crecimiento y fijación de carbono en una plantación de guadua en la zona cafetera de Colombia.”, Recursos Naturales y Ambiente, n.º 61, pp. 86-94, 2010.
[11] A. Castaneda-Mendoza, J. Vargas-Hernandez, A. Gomez-Guerrero, J. I. Valdez-Hernandez, y H. Vaquera-Huerta, “Carbon accumulation in the aboveground biomass of a Bambusa oldhamii plantation“ Agrociencia, vol. 39, n.º 1, pp. 107–116, 2005.
[12] O. Murillo, V. Badilla, y S. Morales, “Método de inventario para plantaciones pequeñas”, Escuela de Ingeniería Forestal, Instituto Tecnológico de Costa Rica. Cartago, Costa Rica, pp. 4–5, 2012.
[13] T. Morales-Pinzón, L. F. Durán, y C. A. Alzate, “Contenido de humedad en guadua rolliza preservada y secada en invernadero.”, Recursos Naturales y Ambiente, n.º 65-66, pp. 45-50, 2012.
[14] I. Widmer, “Caracterización botánica de los bambúes del género Guandua utilizados por el proyecto nacional de bambú en la construcción de viviendas en Costa Rica”, CATIE, 1990.
[15] S. Eggleston, L. Buendia, K. Miwa, T. Ngara, y K. Tanabe, Eds. 2006 IPCC guidelines for national greenhouse gas inventories, vol. 5. Institute for Global Environmental Strategies Hayama, Japan, 2006.
[16] T.-M. Yen, Y.-J. Ji, y J.-S. Lee, “Estimating biomass production and carbon storage for a fast-growing makino bamboo (Phyllostachys makinoi) plant based on the diameter distribution model”, Forest Ecology and Management, vol. 260, n.º 3, pp. 339–344, 2010.
[17] T.-M. Yen y J.-S. Lee, «Comparing aboveground carbon sequestration between moso bamboo (Phyllostachys heterocycla) and China fir (Cunninghamia lanceolata) forests based on the allometric model», Forest Ecology and Management, vol. 261, n.º 6, pp. 995–1002, 2011.
[18] W. Fonseca y M. Rojas, «Acumulación y predicción de biomasa y carbono en plantaciones de bambú en Costa Rica», Ambiente y Desarrollo, vol. 20, n.º 38, pp. 85–98, 2016. doi: 10.11144/Javeriana.ayd20-38.apbc
[19] R. Kaushal et al., “Predictive models for biomass and carbon stock estimation in male bamboo (Dendrocalamus strictus L.) in Doon valley, India”, Acta Ecologica Sinica, vol. 36, n.º 6, pp. 469–476, 2016.
[20] X. Chen, X. Zhang, Y. Zhang, T. Booth, y X. He, «Changes of carbon stocks in bamboo stands in China during 100 years», Forest Ecology and Management, vol. 258, n.º 7, pp. 1489–1496, 2009.
[21] D. Zianis y M. Mencuccini, «On simplifying allometric analyses of forest biomass», Forest Ecology and Management, vol. 187, n.º 2-3, pp. 311–332, 2004.
1 Ingeniero Forestal. Escuela de Biología Instituto Tecnológico de Costa Rica. Costa Rica. Correo electrónico: ebriceno@tec.ac.cr